
| |
Faculty of Science |

|
(Concise description) |
(Names only) |
|
|
|
|
|
|
|
Function & Notes |
| Acetylation | All
All very common in eukaryotes and rare in prokaryotes |
not N,K,R,H,F,W,Y
K |
N-terminus
Anywhere |
- |
One of the most common modifications. Regulates protein stability and protein-DNA interactions (via acetylation of histones) and thus affects transcription. (Acetyl-coA is a required co-factor). Acetylation of N-term occurs in the cytoplasm. |
| Alkylation | Metazoa | C, H, (K) | - |
|
|
| Amidation | Eukaryotes | All | C-terminus | Needs a G after C-terminus on precursor | Amidation of alpha-CO2H occurs in secretory vesicles and granules. |
| Beta-methylthiolation | E.Coli | D | Anywhere |
|
|
| Carbamylation | All | All
K |
N-terminus
Anywhere |
- |
|
| Citrullination | Mammalia | R | Anywhere |
|
|
| C-Mannosylation | Eukaryotes | W | Anywhere |
|
|
| Cysteine sulfenic acid (-SOH) | Metazoa, Bacteria, Plants? | C (side chain) | - |
|
|
| Cysteine sulfinic acid (-SO2H) | Metazoa, Bacteria, Plants? | C (side chain) | - |
|
|
| Cysteine disulfide bond formation | Prokaryotes
Eukaryotes |
C | Anywhere | Cysteine disulfide bond formation occurs in the periplasmic space of bacteria and in the ER of eukaryotes. | |
| Deamidation | All
All |
N
Q |
Anywhere
Anywhere |
Needs to be followed by a G
- |
Possible regulator of protein-ligand and protein-protein interactions. |
| N-acyl diglyceride cysteine (tripalmitate) | Prokaryotes, Archaebacteria, Phages | C | N-terminus after cleavage of signal peptide | Prosite PDOC00013
Consensus pattern: {DERK}(6)-[LIVMFWSTAG](2)-[LIVMFYSTAGCQ]-[AGS]-C
[C is the lipid attachment site] Additional rules: 1) The cysteine must be between positions 15 and 35 of the sequence in consideration. 2) There must be at least one Lys or one Arg in the first seven positions of the sequence. |
|
| Farnesylation | Eukaryotes, Viruses | C | Anywhere |
|
Membrane tethering, reversible |
| Formylation | All | All | N-terminus |
|
|
| Geranyl-geranyl | Eukaryotes, Viruses | C | Anywhere |
|
|
| Gamma-carboxyglutamic acid | Eukaryotes | E | Anywhere |
|
|
| N-linked Glycosylation | Eukaryotes | N, Q | Anywhere |
|
Found in secreted proteins, involved in cell-cell recognition. Core glycosylation of Asn occurs in the ER. Modification of N-glycosyl groups occurs in the Golgi aparatus. |
| O-linked Glycosylation | Eukaryotes | S, T, (N) | Anywhere |
|
O-glycosylation with GalNAc (N-acetylgalactoseamine) occurs in the Golgi apparatus. O-glycosylation with GlcNAc (N-acetylglucoseamine) occurs in the cytoplasm and in the nucleus, is reversible and has regulatory functions. |
| Glucosylation (glycation) | All
All |
All
N, T, K |
N-terminus
Anywhere |
- |
|
| Glutathionylation | Metazoa, Bacteria | C (side chain) | - |
|
|
| Hydroxylation | Eukaryotes | P,K,D,N | Anywhere |
|
Hydroxylation of procollagen Pro and Lys residues occurs in the ER. |
| Methylation | All | All | Anywhere |
|
Regulation of gene expression (via histones methylation). |
| Myristoylation | Eukaryotes, Viruses
Eukaryotes |
G
G K |
N-terminus
N-terminus Anywhere |
|
Myristic acid is co-translationally attached to gly in the seq: N-term met-gly following the cleavage of met. Many signalling proteins are N-myristoylated. Myristoylation is often required for membrane binding of proteins. Myristoylation alone is not enough - a second modificition is normally required to stabilise membrane association. Phosphorylation of the myristoylated protein can destabilise the interaction, releasing the protein from the membrane. Myristoylation occurs in the cytoplasm. |
| N-Octanoate | Mammalia | S: N-Octanoate group linked through an ester bond to a serine residue | Anywhere |
|
|
| Palmitoylation | Eukaryotes | C (S,T,K) | Anywhere
near N- or C- termini or close to transmembrane domains. |
|
Cellular localization and targeting to membrane (stabilises myristoylation). A reversible process. May also be involved in stabilising against protein turnover and/ or delivery to lysosomes. Palmitoylation occurs in the cytoplasm or in the ER. |
| Phosphorylation | Eukaryotes, Viruses
Prokaryotes |
S,T,Y,H,D
S,T,Y,H,C,D |
Anywhere |
|
Reversible, activation/ inactivation of enzyme activity. Regulation of transcription via phosp. of histones. |
| Prenylation | Eukaryotes | C | C-terminus | CAAX box = cysteine, 2x any aliphatic aa, any aa. The nature of X determines whether farnesylation or geranylgeranylion occurs. The addition of the prenyl group to C is followed by proteolytic removal of the AAX sequence and carboxylmethylation of the C-term C. (This step occurs in the endoplasmic reticulum) | Addition of a 15- carbon farnesyl group or a 20- carbon geranylgeranyl moiety. ~ 2% of eukaryotic proteins exist in prenylated form. Prenyl proteins have relatively short half- life (~20 hrs), and their turnover requires removal of the C-prenyl moiety. |
| Sumoylation | Eukaryotes | K | Most SUMO-modified proteins contain the tetrapeptide motif B-K-x-D/E where B is a hydrophobic residue, K is the lysine conjugated to SUMO, x is any amino acid (aa), D or E is an acidic residue. | MW of Sumo-1 is ~ 11 KDa. Related to protein import to the nucleus | |
| Sulfation | Eukaryotes | Y | Anywhere | PROSITE PDOC00003 | Modulation of protein-protein and receptor-ligand interactions. Sulafation occurs in the Golgi apparatus. |
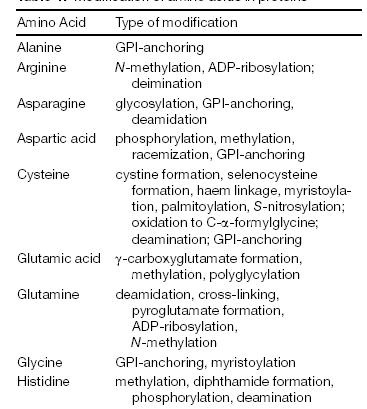 |
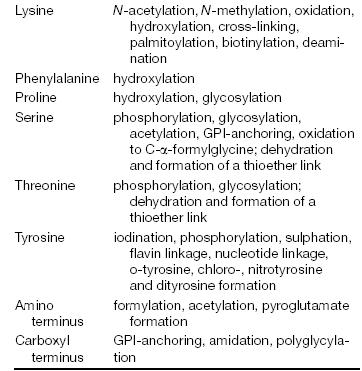 |
Modifications Sorted by Localization: Side Chains vs. Peptide Chain
|
|
|
| proteolytic cleavage, rearrangement, inversion (inteins), N- and C-terminal modification | Phosphorylation, acylation, prenylation, O- and N- glycosylation, nucleosidylation, di sulfide formation, methylation, hypusine, vitamin K dependent carboxylation, hydroxylation, cross-links, etc |
Phosphorylation
in Prokaryotes
Glycosylation
Sumoylation
.
This site is maintained by
Dr. Nurit Doron . Your comments
are most welcome.
Entries since October 2007
Copyright ©, 1997, The Hebrew University of Jerusalem. All Rights Reserved